Meeting report
Bryological symposium
The annual meeting was held on the weekend of 1-2 October in Beaumont Hall situated in the Botanic Gardens of Leicester University. About sixty members and guests attended on the Saturday when the President introduced seven speakers, summaries of whose papers are given here.
D. Glyn Jones (Nature Conservancy Council Aberystwyth): “Aspects of growth and development in Sphagnum cuspidatum.“
The production of innovations on mature Sphagnum shoots was discussed. An innovation is a branch of unlimited growth resembling the parent Sphagnum shoot in that it has a comal tuft and lateral branches in groups, known as fascicles. As Sphagnum shoots die and disintegrate from below, innovations become separated from the parent axis and give rise to new individuals. The frequency with which innovations are produced determines the extent to which Sphagnum is able to exploit its habitat and it is surprising that so little attention has been paid to the factors affecting innovation production the Sphagnum shoot. Results of decapitation experiments suggested that the apex was exerting some sort of control over innovation production, possibly a hormonal type of control.
Since algal infestion of culture solutions containing nitrates hampered experiments with mature shoots, experiments with aseptically-produced primary shoots (shoots from protonemata) were carried out. Originally, the aim of the work described was to investigate the factors affecting the initiation and development of innovations on primary shoots. However, difficulty with interpreting growth of primary shoots in liquid culture led to a study of the developmental morphology of the Sphagnum shoot as a whole.
Shoots grown in balanced inorganic culture solution, with the concentration of ions resembling that of bog water, showed little or no inclination to branch. However. with increased nitrate concentrations branching increased, although these shoots did not resemble mature shoots in that the branches continued growing and comal tufts failed to develop.
Other investigators have failed to grow Sphagnum shoots of normal appearance in liquid culture, unless some organic carbon source was present but no explanation for this requirement was suggested.
The addition of 1% sucrose to low nitrate cultures did not produce Sphagnum shoots of normal appearance. However, addition to high nitrate cultures produced shoots of normal appearance with comal tufts and branches of limited growth, in fascicles.
The production of a great variety of extracellular substances including both simple and complex polysaccharides by a large number of taxonomically diverse algae is now well established.
On testing some inorganic culture solution, containing primary shoots, for soluble carbohydrates by the Anthrone test, a positive result was obtained. The only source of carbohydrate was the Sphagnum shoots themselves. It was concluded that, to grow normally in liquid culture, Sphagnum shoots require sugars to compensate for that lost by leakage. Further evidence for this was that Sphagnum shoots growing into air above inorganic culture medium showed normal growth on the aerial portions.
In a final experiment with S. papillosum, in the presence of high nitrate and high sucrose, 30% more branches were produced than in any other treatment – and only in the presence of high nitrate was a significant proportion of the branches innovations.
It is difficult to relate these experiments directly to the field condition because the concentrations of nitrate found to be effective are relatively so high. It should be remembered, however, that the typical habitat for S. cuspidatum is a shallow depression on the bog surface which is not permanently flooded. During wet periods, therefore, the rain which has a considerably higher concentration of nitrate than bog water, in effect flows through the Sphagnum mat and it is well known that ions are absorbed more efficiently from flowing than from stagnant water. Also, it is likely that the growth-rate of shoots in the lower temperatures of field conditions is slower than in experimental conditions, with the result that more efficient use can be made of the nitrogen absorbed, and furthermore there will be other N2 sources, such as ammonium, in the bog system. So far as sugars are concerned concentrations in excess of 0.01% have been measured in cold water extracts of peat, but because the shoots are exposed for much of the time leakage should be less severe than in water cultures. Also, long unbranched shoots, resembling primary shoots, have been found in permanent pools on ombrotrophic bogs.
The results presented indicated that high levels of nitrogen and sugar together stimulate the development of lateral meristems and, at the same time, limit the period of activity of some, which thus become fascicular branches and promote the activity of others, which become innovations. But so far we have not been able to throw light on the mechanism which controls the type of branch into which a lateral meristem develops.
Dr. D. J. Boatman (Department of Botany, University of Hull): “Experiments on the growth of protonemata of Sphagnum papillosum.“
Protonemata were grown on cellophane discs supported at the surface of a balanced inorganic culture solution in 25 ml beakers (‘Boatman & Lark, 1971). Previous work in which the concentration of the whole nutrient solution and that of individual ions was varied indicated that the growth of protonemata was directly related to the concentration of phosphate and indirectly related to the concentration of calcium. Subsequent investigations revealed that although the pH of the solutions was only slightly affected by autoclaving in the absence of cellophane discs, they were considerably and differentially affected by the presence of discs. Initially the pH of all solutions was about 5.1 but after autoclaving in culture beakers containing cellophane that of the most dilute solutions exceeded 7.0 while that of the most concentrated was about 5.8.
When solutions and the cellophane discs were autoclaved separately the pH of the diluted and undiluted solution was similar even after the culture chambers had been assembled, the range being 5.8 to 6.7. The addition of small quantities of 0.01N hydrochloric acid had a differential effect on the growth of protonemata although the effect on the pH of the various culture solutions was similar. It was concluded that the acid might be having a differential effect on the amounts of metal ions adsorbed on the cellophane discs and this in turn might be affecting the growth of the protonemata.
This hypothesis was tested by treating cellophane discs from the culture chambers with 0.05N nitric acid and determining the amounts of sodium, potassium, magnesium and calcium in the leachate. The amounts of all ions in the leachate decreased with increasing dilution of the culture solution and, at all concentrations, with the addition of hydrochloric acid to the culture solution. Only the amount of calcium in the leachate was affected more by the addition of hydrochloric acid to the culture solution than dilution of the solution as a whole.
Since the addition of acid did not affect the concentration of metal ions in the solutions, diluted or undiluted, it was considered that the protonemata must be depending on adsorbed ions for their growth. Both adsorbed calcium and growth of protonemata are more affected by the addition of acid than dilution of the solution. It is unlikely that the growth of protonemata is directly related to the amount of available calcium but other ions present at low concentrations which are more likely to affect growth, particularly iron, might behave in a similar way to calcium. This has yet to be tested.
Experiments on the growth of protonemata completely immersed in culture solution have also been carried out. No leafy buds developed in inorganic solution but in similar cultures containing glucose at a concentration of 1% almost all of the protonemata produced buds.
Boatman, D. J. & Lark, P. M. (1971). Inorganic nutrition of the protonemata of Sphagnum papillosum Lindb., S. magellanicum Brid. and S. cuspidatum Ehrh. New Phytol. 70, 1053-9.
Dr. M. A. S. Burton (Chelsea College, London University): “Heavy metals in aquatic bryophytes.”
Metal accumulation by terrestrial bryophytes in ore-bearing and polluted habitats is well known, but less information is available on aquatic species. Analysis of bryophytes collected from rivers and streams in a lead-zinc mining area in Wales has demonstrated their capacity to accumulate high concentrations of zinc: up to 7,000 ppm/dry weight in Philonotis fontana; and of lead: up 16,000 ppm/dry weight in Solenostoma crenulatum.
Investigations on the localisation of zinc in aquatic bryophytes showed that the distribution between soluble and insoluble fractions was 10-20% soluble and 80-90% insoluble. There have been similar reports for lead in terrestrial bryophytes, and in algae and grasses zinc has also been shown to be present chiefly in the cell walls. Zinc in the soluble fraction was found to be cationic. A wide range of tolerance to metals is indicated by the occurrence of the bryophytes in waters polluted with up to 50 ppm zinc and 6 ppm. lead and the extent of accumulation was found to reflect the state of metal pollution in the water.
Dr. M. E. Newton (Stalybridge, Cheshire): “Environmental factors controlling sexual reproduction in mosses of the genus Mnium.”
Although Mnium hornum and M. undulatum, are both dioecious, sexual reproduction is rare only in the latter, apparently due to the wide separation of male and female inflorescences. Male plants of M. undulatum are outnumbered by female plants but this alone can not account for the rarity of sporophytes, since most populations are potentially bisexual. The difference appears to be related to male inflorescence development as well. Thus, the regular annual production of male and female inflorescences of M. hornum is determined by an endogenous rhythm, whereas M. undulatum is critically controlled by photoperiod and temperature. Stimuli required by male plants of the latter species are much more exacting than those necessary for female inflorescence production and are such that they are likely to be received by few actively growing male plants in. the field. Spatial separation of male and female gametes of M. undulatum appears, therefore, to be directly related not only to the unequal sex ratio but also to the lower fertility of male than female plants. Since environmental factors determine the frequency of sexual reproduction in M. undulatum, and hence of genetic recombination, they must also control the variability and to some extent the further evolution of the species.
Dr. J. Gorham (Department of Biological Sciences, Portsmouth Polytechnic): “Recent research on lunularic acid.”
Lunularic acid, a bibenzyl compound originally isolated as a dormancy factor in Lunularia cruciata, has been identified together with lunularin in extracts of a wide range of liverwort species. It was not detected in any of the members of the Anthocerotales which were examined, nor in any of the mosses. This finding supports previous chemotaxonomic evidence for the separation of the Anthocerotales from the other orders of liverworts. Contrary to earlier reports, neither lunularic acid nor lunularin could be detected in algae. Traces of lunularic acid, lunularin, 3,4′-dihydroxystilbene and a bound form of lunularic acid, possibly the 3-beta-D-glucopyranoside, were identified in extracts of the roots of Hydrangea macrophylla.
Lunularic acid was found in all parts of Marchantia and Preissia and in sporophytes of Pellia epiphylla. The greatest concentration (more than 600 µg/g fresh weight) was found in the young thallus tips of Conocephalum conicum grown in continuous light. In general the quantities of lunularic acid present increased at higher light intensities and decreased in older tissues. Members of the Jungermanniales contained smaller quantities of lunularic acid (between 1 and 50 µg/g fresh weight). When thallose liverworts were grown in different daylengths, in which a basic 8 hr photoperiod was extended by light of the same intensity, both fresh weight increase and lunularic acid content were greater in continuous light. In these conditions either lunularic acid was not inhibitory or the inhibition was overcome by the products of photosynthesis.
When tested in Marchantia and Lunularia gemmaling assays and the cress root growth test, lunularic acid was not found to be more effective an inhibitor than a wide range of similar compounds. Indeed, lunularin was slightly more active than lunularic acid. No correlation between structure and inhibitory activity was observed for a number of analogues of lunularic acid.
Attempts to relate the inhibitory activity of lunularic acid and its analogues to their ability to modify indol-3ylacetic acid oxidase (IAA-oxidase) activity were frustrated by the ambiguous nature of results obtained from in vitro experiments. Both stimulation and inhibition of the lAA-oxidase activity of horseradish peroxidase were observed, depending on the nature and concentration of other cofactors. In the presence of 2,4-dichlorophenol, polyphenols were generally inhibitory to IAA-oxidase activity whereas carboxylic acids, monophenols and non-phenolic compounds had little effect. Whilst the IAA-oxidase modifying effect of strong acid fractions of extracts of Conocephalum was attributable to lunularic acid, a stronger modifier, which had the properties of a phenolic glycoside, was found in aqueous extracts. Inhibition of glucose-6-phosphate dehydrogenase activity by lunularic acid was also observed.
The proposed biosynthesis of lunularic acid and lunularin via the phenyl-propanoid pathway was supported by the demonstration of the enzymes phenyl-alanine ammonia lyase, cinnamic acid-4-hydroxylase and lunularic acid decarboxylase in Conocephalum, and by tracer studies. 14C-phenylalanine was more readily incorporated into lunularic acid than was tyrosine. Over 16 hours, 12.5% of phenylalanine fed to Conocephalum was converted to lunularic acid. Time-course and pulse-labelling experiments showed that radioactivity derived from 14C-phenylalanine remained high in lunularic acid and lunularin over a period of 144 hours, whereas radioactivity in 14C-lunularic acid declined rapidly when it was fed to Conocephalum. An analysis of material which had been incubated with 14C-lunularic acid revealed that most of the radioactivity had been incorporated into an insoluble residue which was resistant to attack by lytic enzymes and solvents.
The role of lunularic acid as an endogenous dormancy factor in all liverworts is not established. The compartmentation of lunularic acid metabolism within cells of liverworts renders interpretation of extraction and assay experiments hazardous.
Dr. M. C. F. Proctor (Department of Biological Sciences, The University, Exeter): “Structure, function and environment in some poikilohydric bryophytes.”
Bryophytes share the poikilohydric habit with many other plants and animals, e.g. many bacteria, lichens, seeds, nematodes and rotifers. Tolerance of desiccation is a prerequisite for all, and features of response to desiccation may be common to widely differing organisms.
Behaviour in relation to water during moist periods is also important in the biology of poikilohydric bryophytes. Species of continuously moist habitats (e.g. Pellia epiphylla, Hookeria lucens) reach maximum photosynthesis only at water contents in the region of 500% to 1000% of dry weight; respiration is little affected until very much lower water contents are reached. In such species as Tortula intermedia and Anomodon viticulosus photosynthesis and respiration show a much more nearly parallel response to water content, but with a strongly marked decline in photosynthesis at high water contents due to the superincumbent water interfering with diffusion of carbon dioxide. It is suggested that strongly poikilohydric species possess adaptations that tend to regulate the distribution of water to the photosynthesising shoots and maximise the time at near-optimal water content. The papillose cells of Tortula intermedia provide a system of capillary spaces covering the surface. The volume of the spaces is too small for significant water storage in an exposed situation, but calculation shows that they provide a conducting system adequate to balance high rates of evaporation in the field as long as water is freely available at the leaf base. Thuidium tamariscinum combines papillose abaxial leaf surfaces and the conducting ‘wick’ of paraphyllia on the stems with storage in the non-papillose concavities of the branch leaves; Cololejeunea rossettiana shows a somewhat similar combination of features. In bryophytes with smooth leaf surfaces conduction through the cell walls is likely to be important, and this may be a major factor in the evolution of the thick cell walls of many species of dry habitats. At low wind speed, low radiation income and high humidity (as in sheltered woods) evaporation rates may be very much lower, and local storage of water becomes a viable adaptation for prolonging periods of near-optimal cell water content. Structures such as the “water sacs” formed by the postical lobes of the small Lejeuneaceae may function in this way.
Dr. S. R. Edwards (The Museum, Manchester): “Taxonomic implications of cell patterns in haplolepidous moss peristomes.”
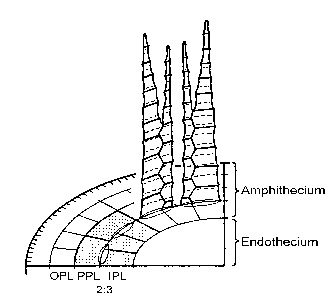 Since Philibert divided the arthrodontous mosses (Bryopsida) into the Haplolepidae and Diplolepidae in 1884, bryologists have questioned whether these groups are natural. The problem has been confused by taxonomically misplaced families, and also by the misassumption that haplolepidy means a single peristome and that diplolepidy means a double peristome.
Since Philibert divided the arthrodontous mosses (Bryopsida) into the Haplolepidae and Diplolepidae in 1884, bryologists have questioned whether these groups are natural. The problem has been confused by taxonomically misplaced families, and also by the misassumption that haplolepidy means a single peristome and that diplolepidy means a double peristome.
The haplolepidous peristome (and also the inner peristome of the Diplolepidae) is formed from thickenings of the periclinal wall-pairs common to the innermost two cell layers of the amphithecium (working outwards, the inner three amphithecial layers have been termed by Blomquist and Robertson the Inner, Primary and Outer Peristomial Layers, or IPL., PPL., OPL in Fig. 1 – a stereogram of a haplolepidous peristome loosely based on Dicranella heteromalla.). In the Bryopsida there are almost always 16 columns of cells in the PPL, but only on the Haplolepidae is each pair of columns (or pair of teeth) faced with three columns of IPL cells; in the Diplolepidae any number except three may be found. Thus haplolepidous teeth are alternately left and right handed, each being faced by 1½ IPL cell-columns; in the Diplolepidae all teeth are generally symmetrical.
Although a 16:24 PPL:IPL ratio had previously been noted in transverse sections of haplolepidous moss capsules by Evans and Hooker, Kreulen, and Mueller, it has never been recognised in mature teeth where ironically it is most easily seen. In section the 16:24 ratio usually becomes approximate owing to irregularities in peristome development and also to the limited vertical extent of the pattern; but when even very reduced haplolepidous peristomes are viewed from the inside it is generally possible to detect a 2:3 PPL:IPL ratio. Moreover, the above authors have each reported the pattern only in one species or in one case two species in one family, and none has suggested it as an essential haplolepidous character.
Thus a survey was conducted of over 60 arthrodontous species to see i) whether the 2:3 ratio was universal to all the five haplolepidous orders (Dicranales, Fissidentales, Pottiales, Syrrhopodontales and Grimmiales), and ii) whether it was absent from the remaining eight diplolepidous orders. The PPL:IPL ratio of 2:3 was indeed found throughout all five haplolepidous orders, and the only “diplolepidous” species found to shew this pattern (such as Ptychomitrium and Glyphomitrium) are clearly misplaced. It is thus concluded that the Haplolepidae form a natural monophyletic group characterised by a PPL:IPL ratio of 2:3 except in species where the peristome is very degenerate or secondarily aberrant. All Encalypta species examined shew an OPL:PPL:IPL ratio of 4:2:4. which together with the frequent development of an outer peristome weakens any suggestion that the Encalyptales form a link between the Haplolepidae and the Diplolepidae. Also, characters distinguishing the Dicranales from the Grimmiales were discussed, and it was suggested that the Seligeriaceae is ill-placed in the Dicranales. In Splachnum ampullaceum, 16 vertical ducts were noticed which communicated between the air-sac and the PPL where PPL/IPL walls ruptured upon dehiscence.
Conversazione
| The A. G. M. was held after tea. In the evening members were generously provided with a reception at Beaumont Hall where the following were displayed: | |
| Dr. H. J. B. Birks: | “Photographs of some British Hepatics.” |
| Dr. K. J. Adams: | “Publications of the British Bryological Society. “ |
| Dr. S. R. Edwards: | “Stereograrns of moss peristomes.” |
| Drs. H. L. K. & M. P. Whitehouse: | “A nest of Carder Bees, Bombus agrorum, composed of Eurhynchium praelongum.“ |
The Society is most grateful to Dr. C. A. Stace (Department of Botany, University of Leicester) who acted as local secretary not only for the organisation of this meeting, but also for making arrangements for members to stay at Beaumont Hall over the weekend.
J.G. Duckett
Field Meeting – Rutland and South Lincolnshire
On 2 October, members explored several localities in Rutland (v.-c. 55) and South Lincolnshire (v.-c. 53) to the north of Stamford. The convoy of cars was indicative of the interest engendered in the autumn meeting as a whole and not necessarily the field trip. But it was pleasing to see such a good turn-out to study bryophytes in Lincolnshire – a county not hitherto visited by the B. B. S.
Ketton limestone quarry (43/974057, v.-c. 55) furnished a useful list of over 50 bryophytes, the most interesting of which were Aloina aloides var. aloides and Rhynchostegiella tenella.
After refreshment at “The George”, Stamford, bryology was renewed with full vigour. At Holywell (53/006161, v.-c. 53) one of the few remaining areas of limestone grassland hereabouts was investigated. Unfortunately a change in management of the area had radically changed the flora in recent years: it proved bryologically and lichenologically poor. and the single remaining plant of Thesium humifusum was located by Miss Conolly in a manner best described as “buried treasure pacing”. A range of substrates was investigated, including a streamside, trees, logs and walls, but the most profitable habitats were located in an abandoned garden (53/004163). Dicranella schreberana* found on moist clay by the stream was amongst the notable finds which also included Tortula laevipila on a log, and Neckera crispa which has been rarely seen in the county this century. The poor hepatic flora here was typical of all the sites visited on the excursion.
[* = new vice-county record.]
Clipsham quarry (43/081154, v.-c. 55) provided the most interesting bryophyte flora. Thuidium abietinum on a roadside, Brachythecium glareosum on a limestone bank, and Gyroweisia tenuis c. spor. in limestone crevices were particularly handsome. The epiphytic flora was relatively good here, and a Prunus thicket supported Bryum flaccidum*, Orthotrichum affine and Radula complanata. A form of Bryum bicolor with rhizoid tubers was collected by Mr Long.
Lincolnshire Gate (53/002148. v.-c. 53) contrasted markedly with the other sites visited; here one of the few pockets of sand in the region is found. Unfortunately, by this time the members were strung out over the countryside (many of them still crawling around Clipsham quarry), and the site did not receive the full attention it deserved. Mr Wanstall displayed his taxonomic skills in a failing light – large areas of the sand being in various stages of colonisation by Polytrichum spp. with particularly fine patches of P. piliferum and P. urnigerum.
M R D Seaward